
Discovery and Basic Research - Biology
Category: Late Breaking Poster Abstract
.jpg "Sarah Cash, BS photo")
Sarah Cash, BS
PhD Candidate
University of Toronto
Toronto, Ontario, Canada
Sarah Cash, BS
PhD Candidate
University of Toronto
Toronto, Ontario, Canada
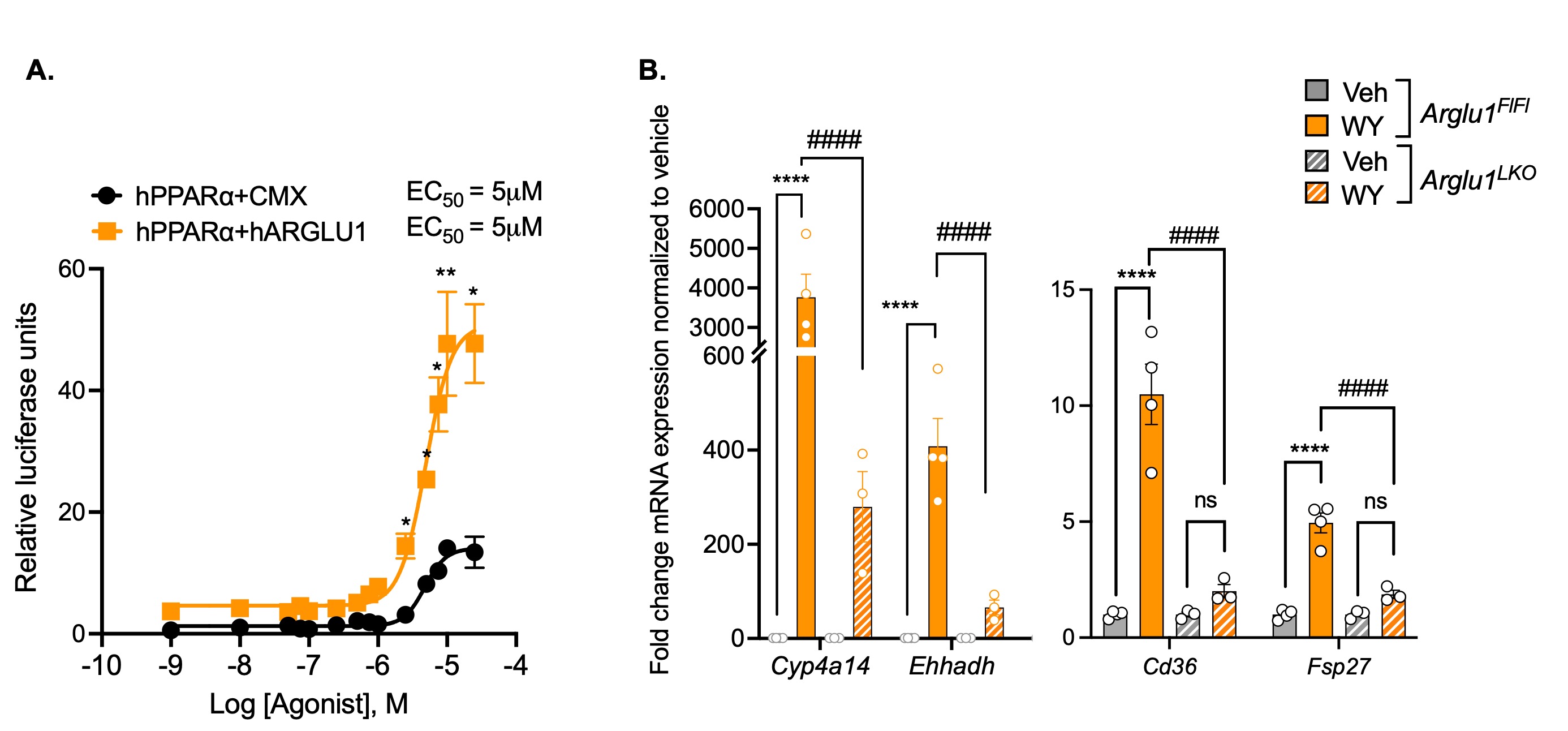
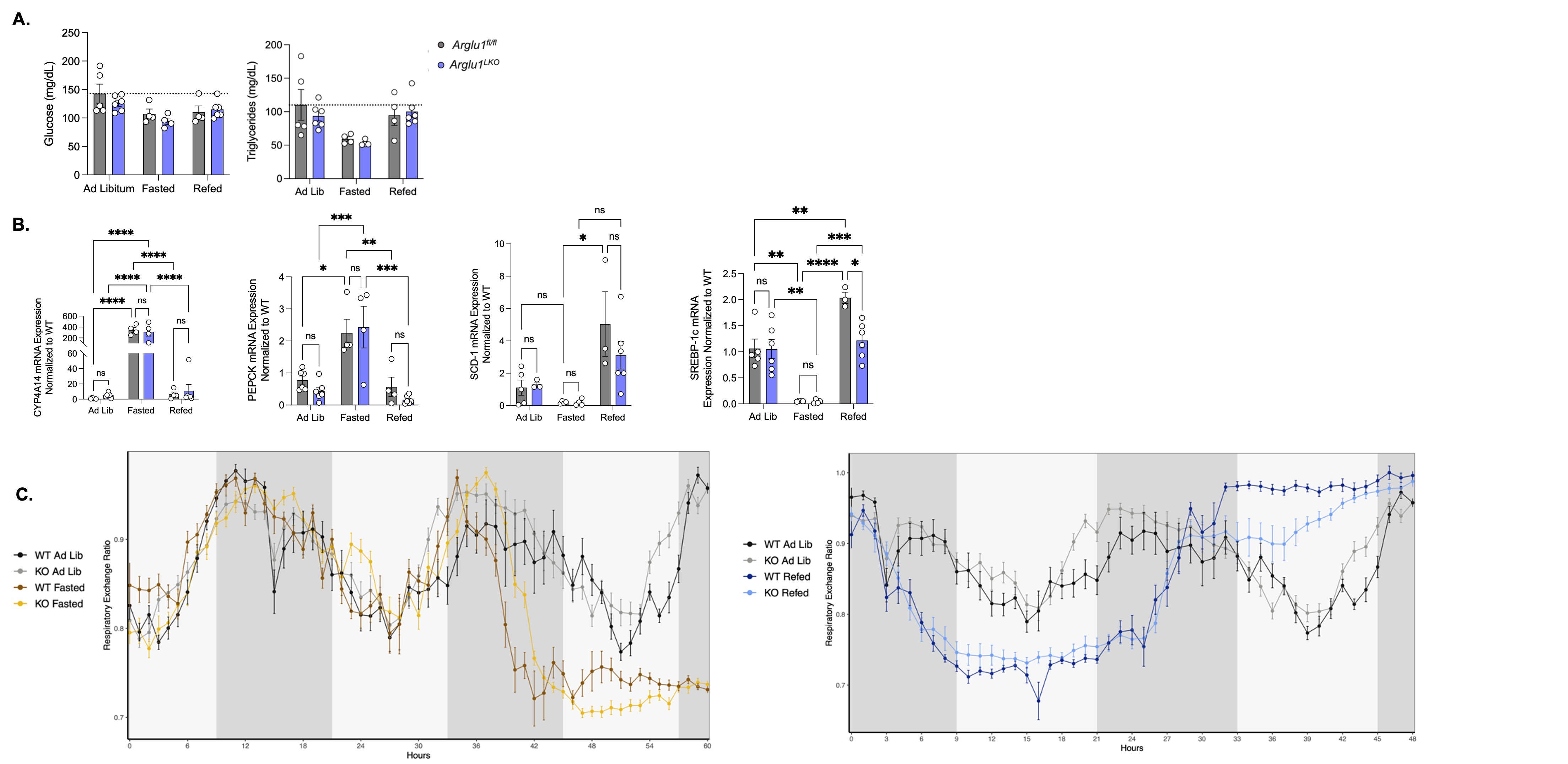
.jpg)