
Poster Session C
Genetics, genomics and proteomics
Katherine Owen, PhD
RILITE
Charlottesville, VA, United States
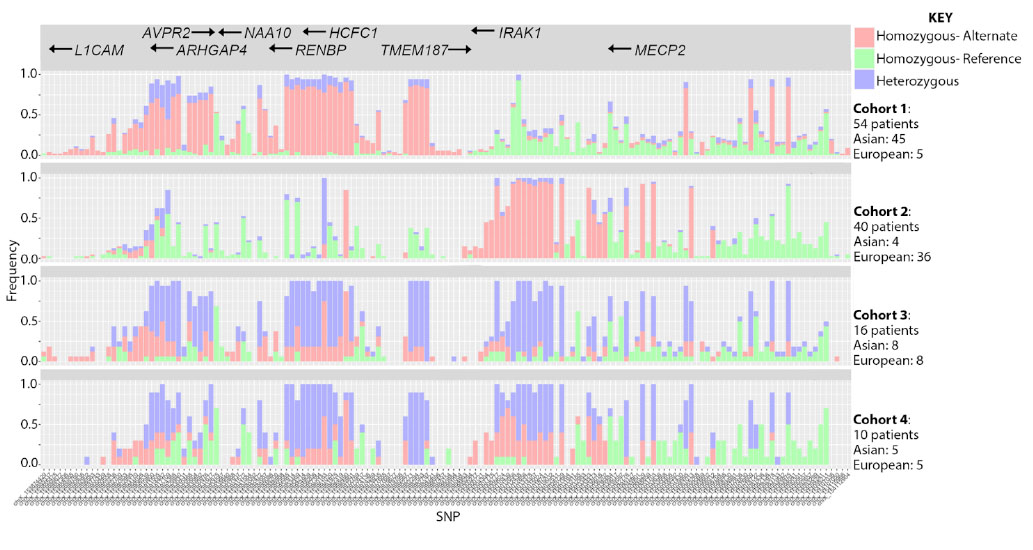 Figure 1: SNP expression, detection and cohort separation annotated with size and ancestral make-up of 4 independent cohorts determined by t-SNE.
Figure 1: SNP expression, detection and cohort separation annotated with size and ancestral make-up of 4 independent cohorts determined by t-SNE.