
Abstract Session
Systemic lupus erythematosus (SLE)
Richard Moore, BSc
Cedars-Sinai
Los Angeles, CA, United States
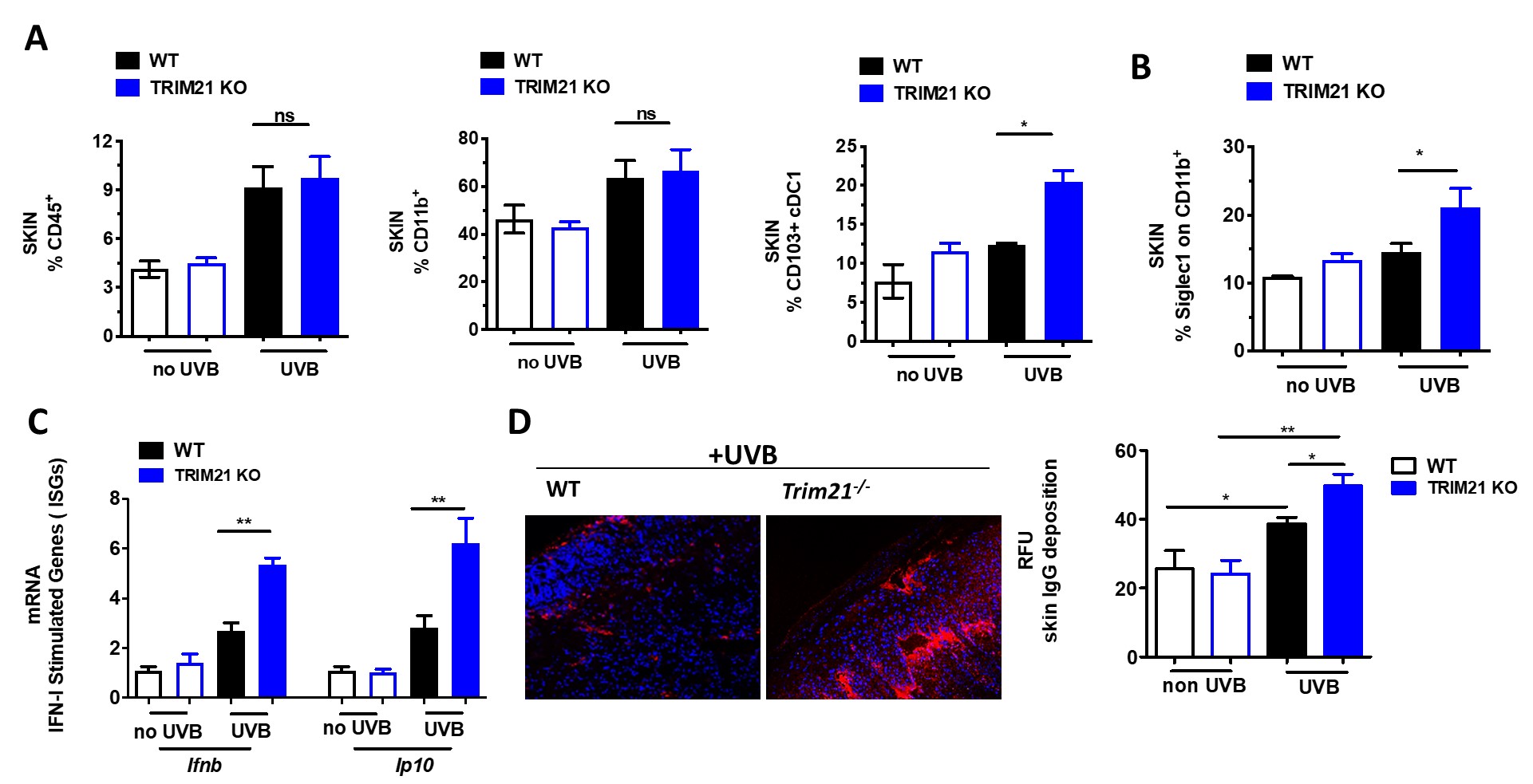 Figure 1. C57BL/6 Trim21+/+ (WT) and Trim21-/- (TRIM21 KO) mice were exposed to UVB irradiation. (A) Flow cytometry analysis following the final UVB dose showing the percentage of infiltrating immune cells (CD45+) , myeloid cells (CD11b+), dendritic cells (CD45+CD11C+MHCII+CD103+). (B) The percentage of Siglec1, an IFN-inducible protein, on CD11b+ skin infiltrating cells. (C) Cutaneous expression of mRNA for IFN inducible genes (ISGs) genes following UVB injury, as determined by qPCR. Fold change was calculated against nonirradiated mice, using the 2−ΔΔCt method. Data are compiled from at least 3 independent experiments. (D) IgG deposition in UVB induced skin lesion. A representative pictures and quantification analysis. Values are the mean ± SD. *p < 0.05; versus no UV irradiation, One-Way ANOVA with Tukey’s post-hoc test *p < 0.05, **p < 0.01.
Figure 1. C57BL/6 Trim21+/+ (WT) and Trim21-/- (TRIM21 KO) mice were exposed to UVB irradiation. (A) Flow cytometry analysis following the final UVB dose showing the percentage of infiltrating immune cells (CD45+) , myeloid cells (CD11b+), dendritic cells (CD45+CD11C+MHCII+CD103+). (B) The percentage of Siglec1, an IFN-inducible protein, on CD11b+ skin infiltrating cells. (C) Cutaneous expression of mRNA for IFN inducible genes (ISGs) genes following UVB injury, as determined by qPCR. Fold change was calculated against nonirradiated mice, using the 2−ΔΔCt method. Data are compiled from at least 3 independent experiments. (D) IgG deposition in UVB induced skin lesion. A representative pictures and quantification analysis. Values are the mean ± SD. *p < 0.05; versus no UV irradiation, One-Way ANOVA with Tukey’s post-hoc test *p < 0.05, **p < 0.01. .jpg) Figure 2. C57BL/6 Trim21+/+ (WT) and Trim21-/- (TRIM21 KO) mice were exposed to UVB irradiation. (A) Compared to WT mice, Trim21-/- mice post-UVB exposure had higher circulating monocytes and increased inflammatory markers CCR2 and Siglec1 in Ly6Chi and CD11b+ cells, respectively. (B) Inflammatory cytokines post-UVB determined by Legendflex assay. (C) FACS analysis of spleens following UVB exposure. n = 5-6 per group. All data represent mean ± SD. Statistical significance was determined using One-Way ANOVA with Tukey’s post-hoc test . *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 2. C57BL/6 Trim21+/+ (WT) and Trim21-/- (TRIM21 KO) mice were exposed to UVB irradiation. (A) Compared to WT mice, Trim21-/- mice post-UVB exposure had higher circulating monocytes and increased inflammatory markers CCR2 and Siglec1 in Ly6Chi and CD11b+ cells, respectively. (B) Inflammatory cytokines post-UVB determined by Legendflex assay. (C) FACS analysis of spleens following UVB exposure. n = 5-6 per group. All data represent mean ± SD. Statistical significance was determined using One-Way ANOVA with Tukey’s post-hoc test . *p < 0.05, **p < 0.01, ***p < 0.001.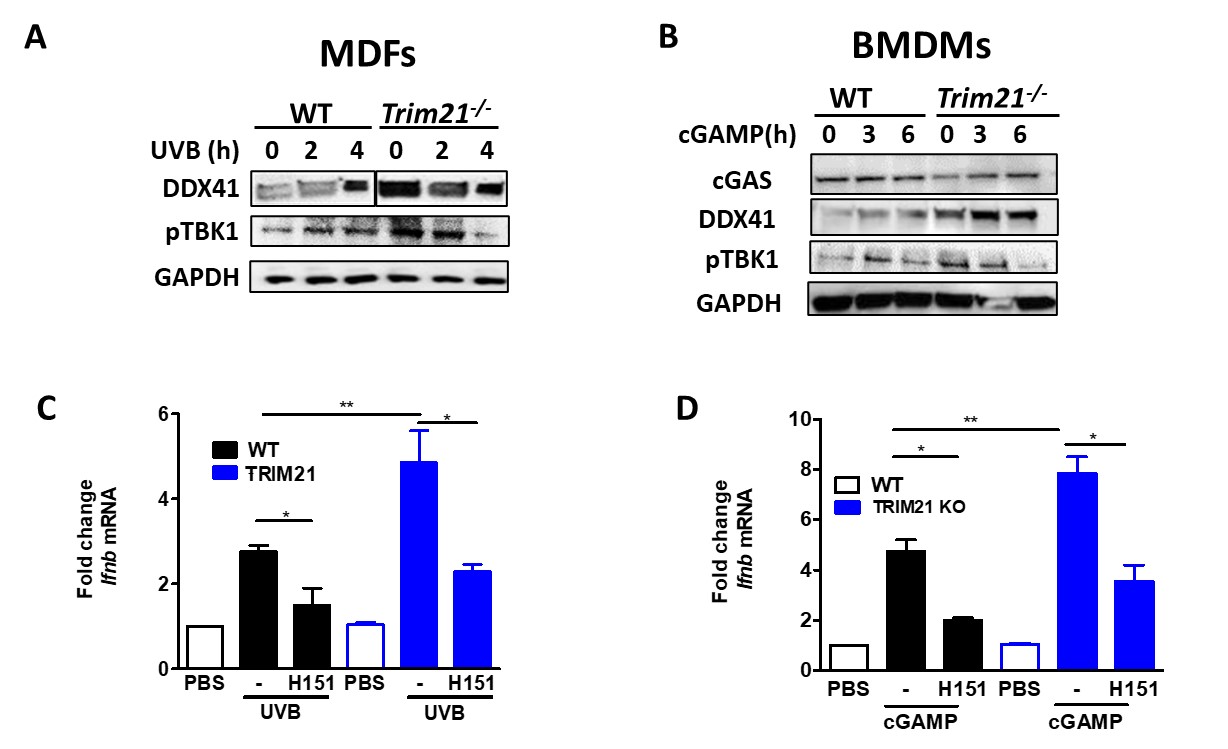 Figure 3. Mouse Dermal Fibroblasts (MDFs) and bone marrow derived macrophages (BMDM) derived from WT and Trim21-/- mice were stimulated with UVB or cGAMP for indicated hours. (A-B) The levels of DDX41, cGAS and pTBK1 were determined by immunoblotting. (C-D) The expression of Ifnb mRNA was determined by qPCR. All data represent mean ± SD. Statistical significance was determined using t-test in A; One-Way ANOVA with Tukey’s post-hoc test in B . *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 3. Mouse Dermal Fibroblasts (MDFs) and bone marrow derived macrophages (BMDM) derived from WT and Trim21-/- mice were stimulated with UVB or cGAMP for indicated hours. (A-B) The levels of DDX41, cGAS and pTBK1 were determined by immunoblotting. (C-D) The expression of Ifnb mRNA was determined by qPCR. All data represent mean ± SD. Statistical significance was determined using t-test in A; One-Way ANOVA with Tukey’s post-hoc test in B . *p < 0.05, **p < 0.01, ***p < 0.001.