
Ignite Talk
Marc Scherlinger, MD, PhD
Beth Israel Deaconess Medical Center Harvard University
Strasbourg, France
Disclosure: Disclosure information not submitted.
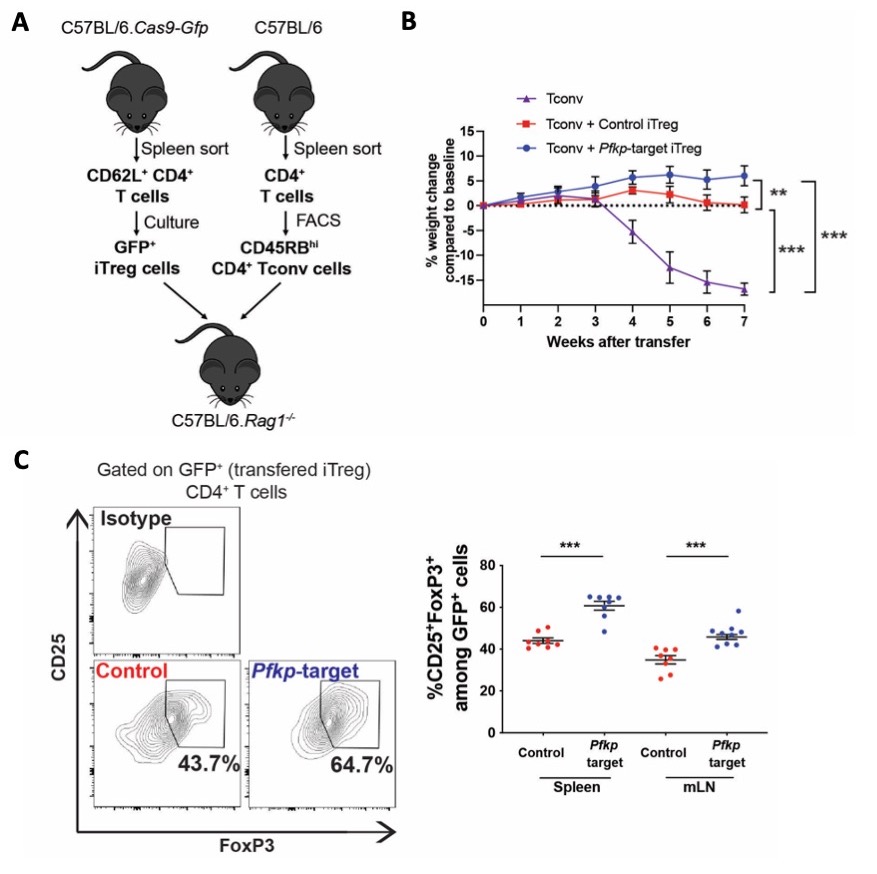 Figure 1: PFKP modulation improves the iTreg cell in vivo function and stability in the adoptive transfer colitis model.
Figure 1: PFKP modulation improves the iTreg cell in vivo function and stability in the adoptive transfer colitis model.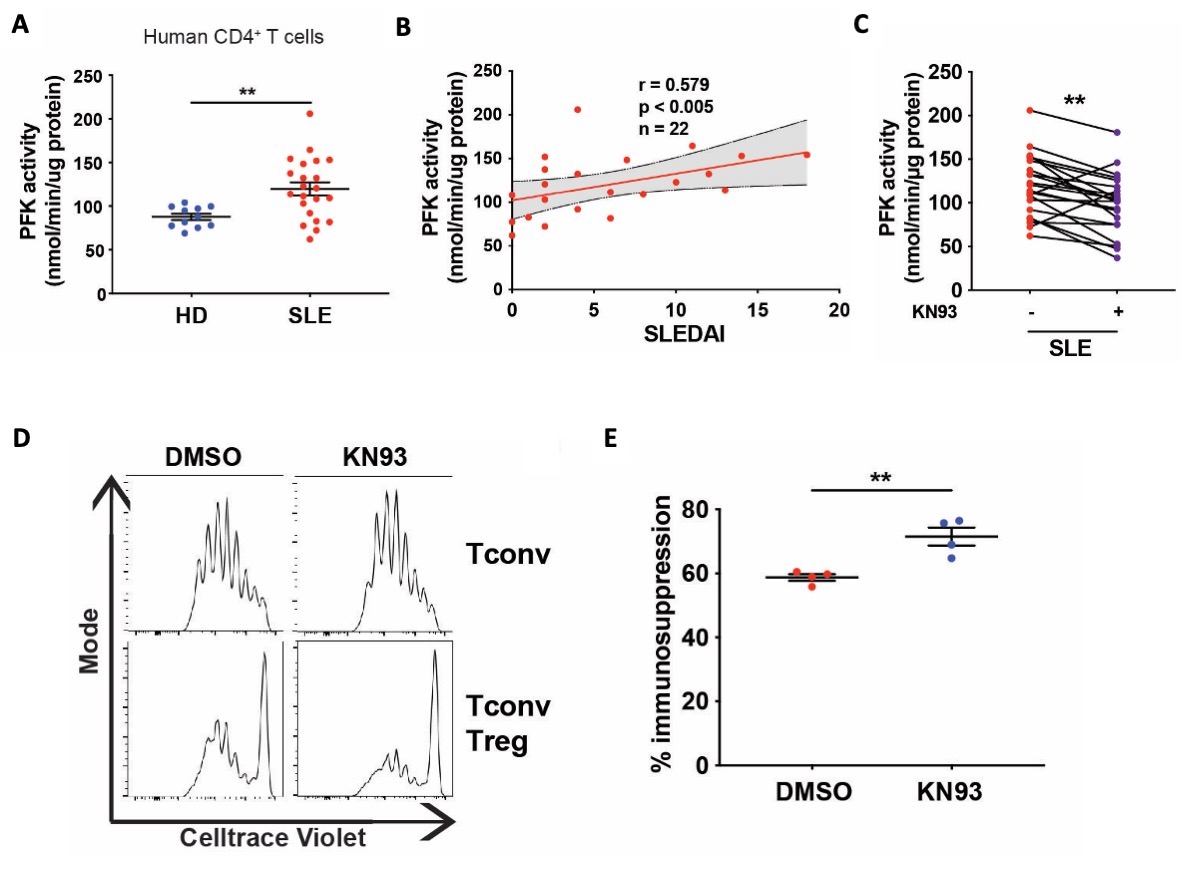 Figure 2: CaMK4 regulates PFKP activity in SLE CD4+ T cells and impairs human Treg cell immunosuppressive functions.
Figure 2: CaMK4 regulates PFKP activity in SLE CD4+ T cells and impairs human Treg cell immunosuppressive functions. 