
Abstract Session
Rheumatoid arthritis (RA)

Melanie Smith, MD, PhD
Hospital for Special Surgery
New York, NY, United States
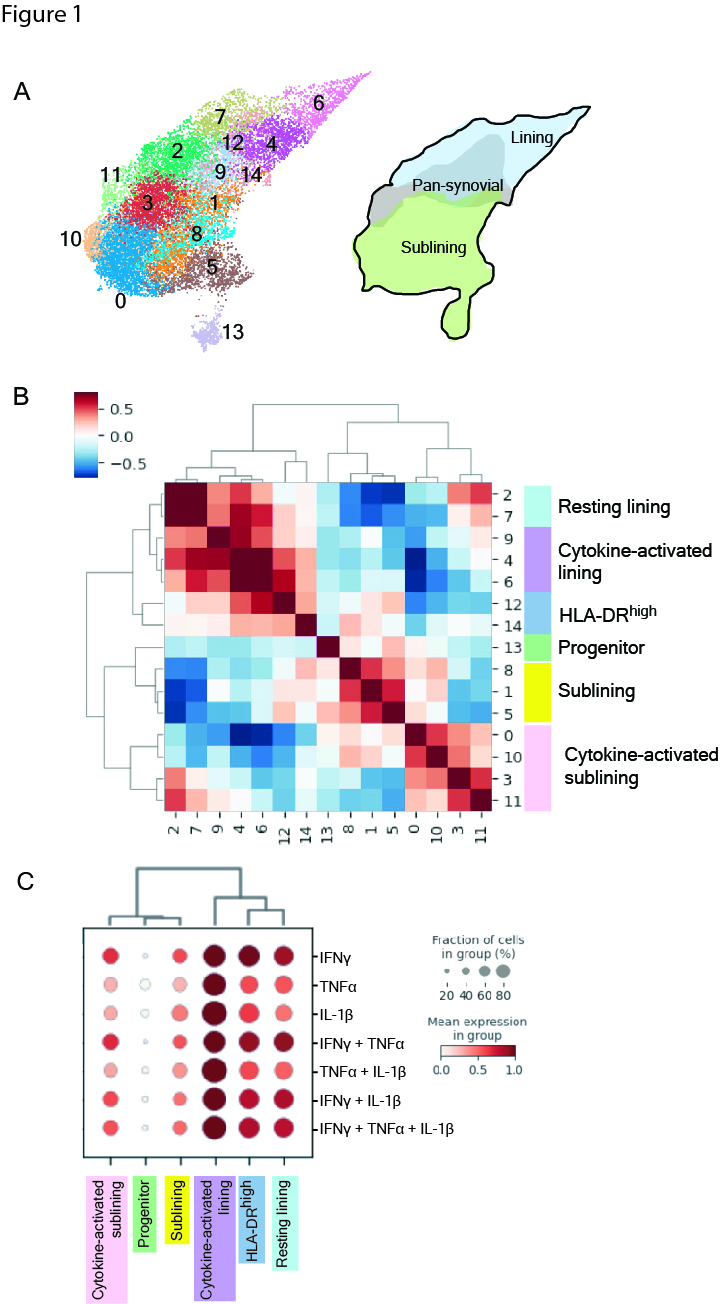 Figure 1. FLS in the inflamed RA synovium exist in distinct states driven by cytokine signaling.
Figure 1. FLS in the inflamed RA synovium exist in distinct states driven by cytokine signaling. .jpg) Figure 3. FLS states and cytokine signaling are spatially constrained within the synovium.
Figure 3. FLS states and cytokine signaling are spatially constrained within the synovium.