
Poster Session D
Immunobiology
Patricia Pontes Aires, MD
Universidade Federal de Sao Paulo
São Paulo, S�o Paulo, Brazil
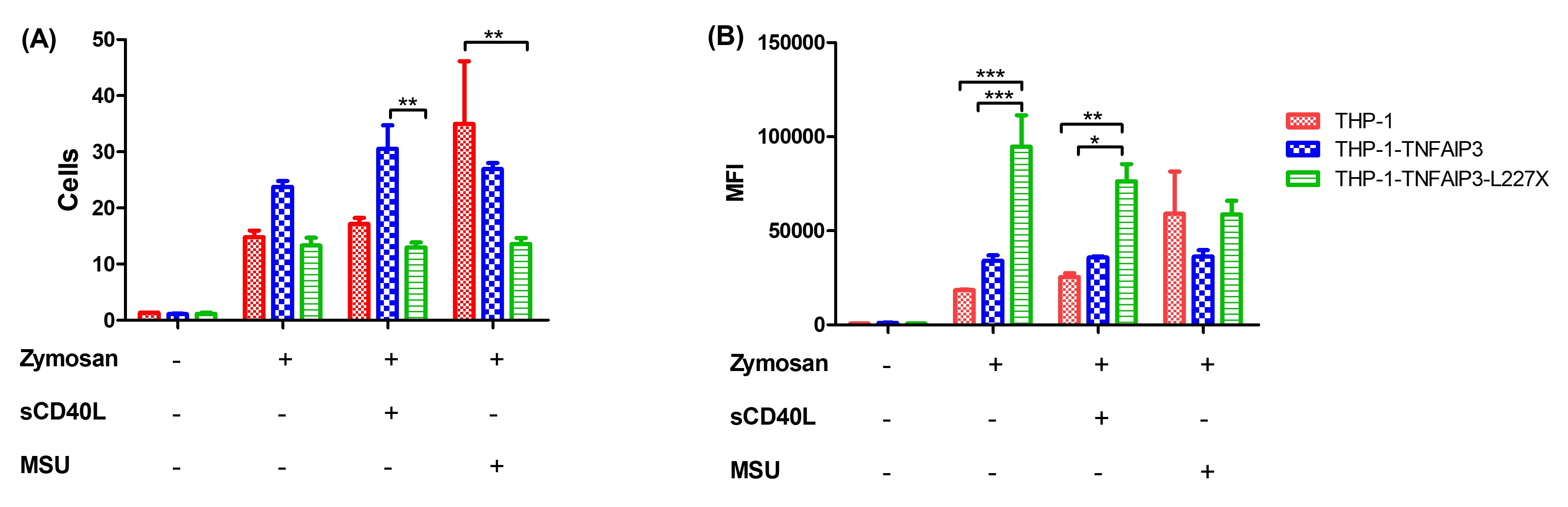 Figure 1: phagocytosis of Alexa-Fluor 594-tagged zymosan particles by THP-1 cells. Comparison of THP-1, THP-1-TNFAIP3-WT and THP-1-TNFAIP3-L227X cells, in triplicate, using two-way ANOVA. Results indicate higher phagocytic rates for THP-1-TNFAIP3-L227X at baseline and upon stimulation with sCD40L * p < 0.05; ** p < 0.01; *** p < 0.001
Figure 1: phagocytosis of Alexa-Fluor 594-tagged zymosan particles by THP-1 cells. Comparison of THP-1, THP-1-TNFAIP3-WT and THP-1-TNFAIP3-L227X cells, in triplicate, using two-way ANOVA. Results indicate higher phagocytic rates for THP-1-TNFAIP3-L227X at baseline and upon stimulation with sCD40L * p < 0.05; ** p < 0.01; *** p < 0.001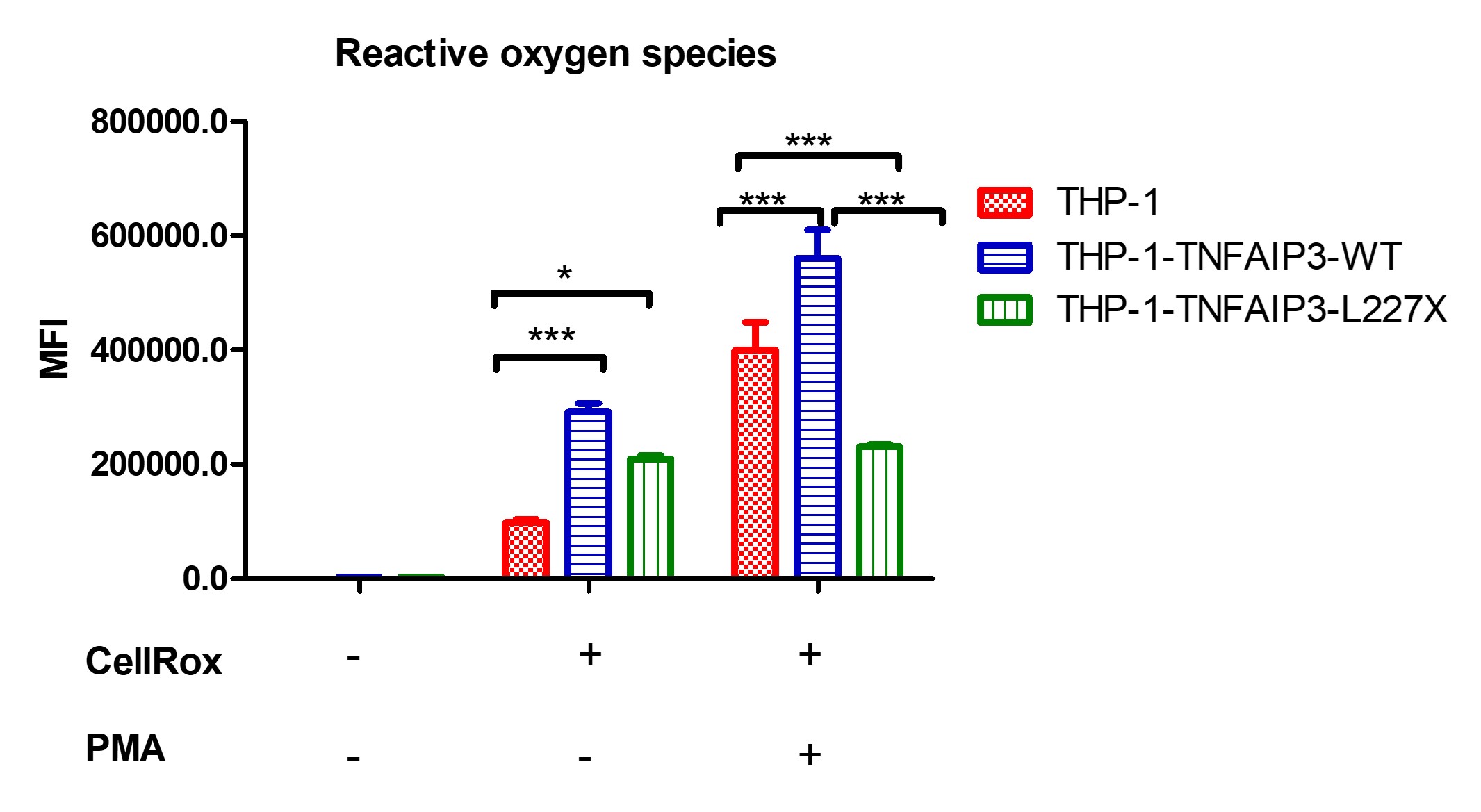 Figure 2: production of ROS by THP-1 cells, before and after PMA stimulus. Comparison of THP-1, THP-1-TNFAIP3-WT and THP-1-TNFAIP3-L227X cells, in triplicate, using two-way ANOVA. Results indicate higher oxidative burst in THP-1-TNFAIP3-WT cells and unresponsiveness of THP-1-TNFAIP3-L227X, which could indicate a state of cell exhaustion. * p < 0.05; *** p < 0.001
Figure 2: production of ROS by THP-1 cells, before and after PMA stimulus. Comparison of THP-1, THP-1-TNFAIP3-WT and THP-1-TNFAIP3-L227X cells, in triplicate, using two-way ANOVA. Results indicate higher oxidative burst in THP-1-TNFAIP3-WT cells and unresponsiveness of THP-1-TNFAIP3-L227X, which could indicate a state of cell exhaustion. * p < 0.05; *** p < 0.001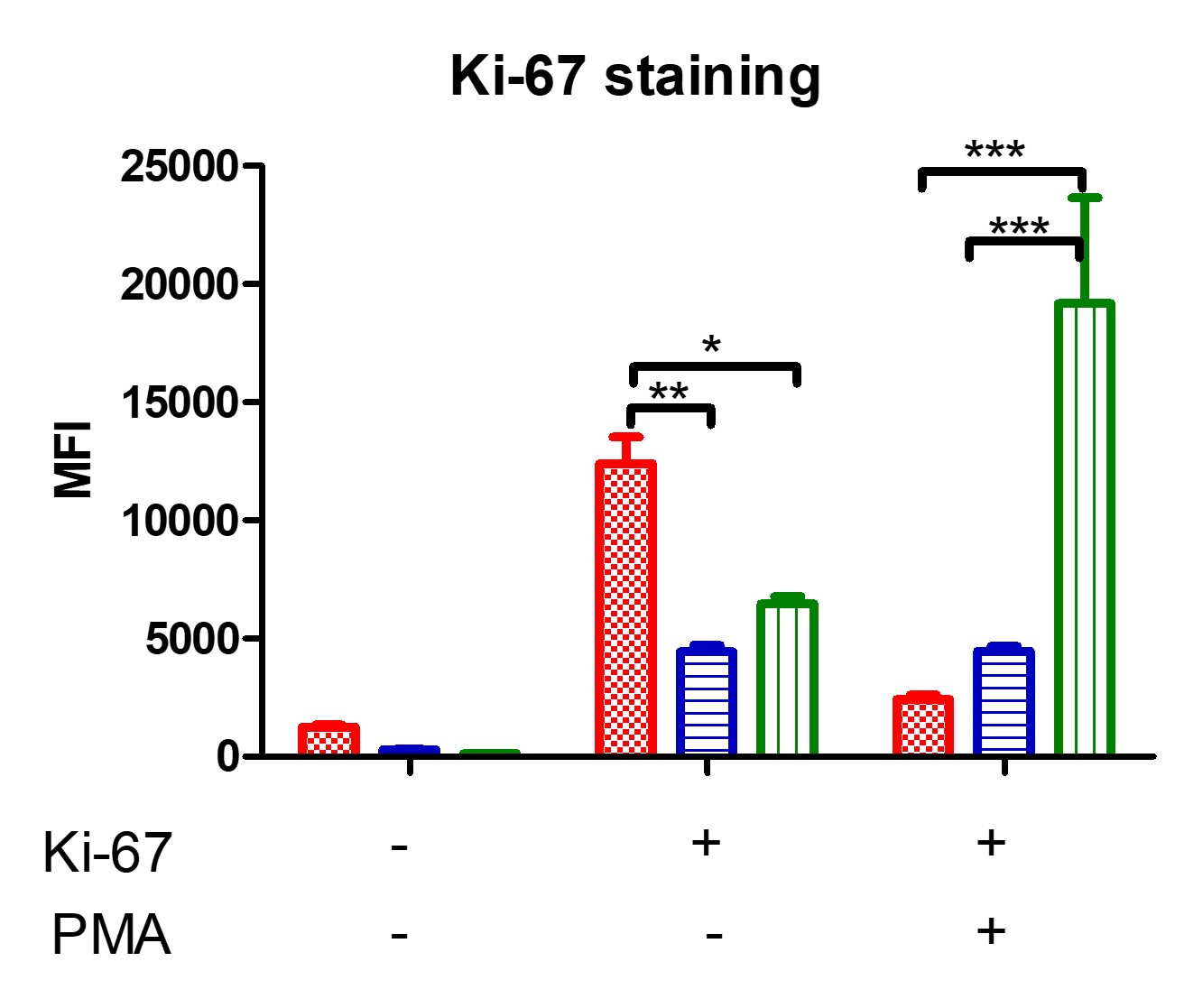 Figure 3: analysis of THP-1 cell proliferation using flow cytometry, through Ki-67 staining, before and after an 18-hour incubation with PMA. Comparison of THP-1, THP-1-TNFAIP3-WT and THP-1-TNFAIP3-L227X cells, in triplicate, using two-way ANOVA. Results show higher proliferation rates for THP-1-TNFAIP3-L227X and low proliferation rates for THP-1-TNFAIP3-WT. * p < 0.05; ** p < 0.01; *** p < 0.001
Figure 3: analysis of THP-1 cell proliferation using flow cytometry, through Ki-67 staining, before and after an 18-hour incubation with PMA. Comparison of THP-1, THP-1-TNFAIP3-WT and THP-1-TNFAIP3-L227X cells, in triplicate, using two-way ANOVA. Results show higher proliferation rates for THP-1-TNFAIP3-L227X and low proliferation rates for THP-1-TNFAIP3-WT. * p < 0.05; ** p < 0.01; *** p < 0.001