
Abstract Session
Thomas Vazquez, MD
University of Pennsylvania
Philadelphia, PA, United States
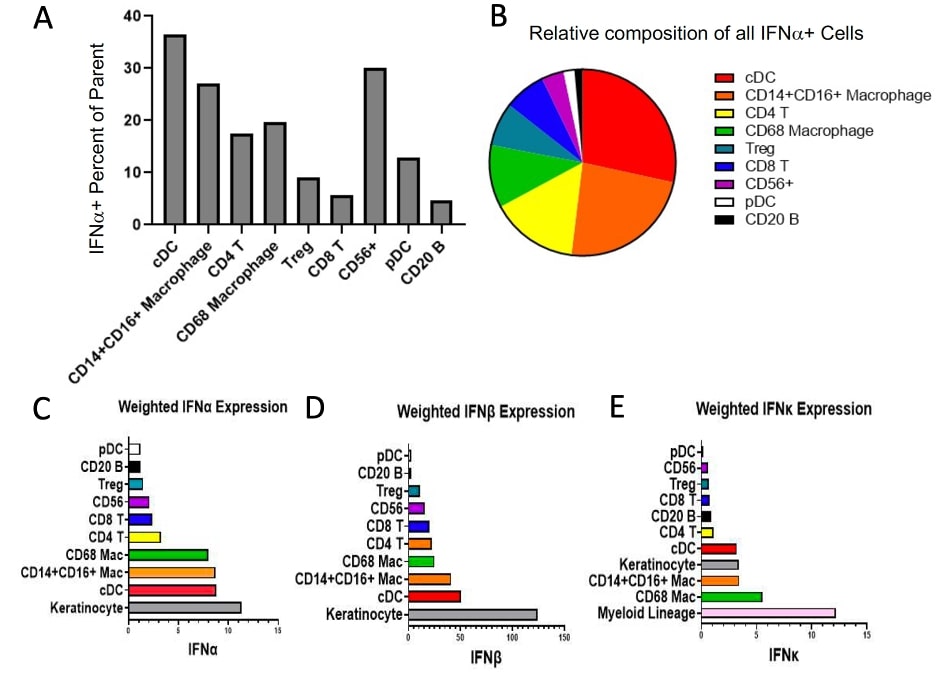 A) IFNα+ Percent of Parent for cell types identified in CLE skin shown in order of descending absolute contribution of IFNα+ cell counts in CLE. 12.83% of pDCs are positive for IFNα. B) Composition of IFNα+ cells (absolute counts) across all aggregated CLE biopsies. C) Weighted (cell count x MPI) IFNα expression. D) Weighted IFNβ expression. E) Weighted IFNκ expression with myeloid lineage consisting of CD68 Mac, CD14+CD16+ Mac, and cDCs revealing dominant IFN-1 contribution from myeloid cells followed by keratinocytes.
A) IFNα+ Percent of Parent for cell types identified in CLE skin shown in order of descending absolute contribution of IFNα+ cell counts in CLE. 12.83% of pDCs are positive for IFNα. B) Composition of IFNα+ cells (absolute counts) across all aggregated CLE biopsies. C) Weighted (cell count x MPI) IFNα expression. D) Weighted IFNβ expression. E) Weighted IFNκ expression with myeloid lineage consisting of CD68 Mac, CD14+CD16+ Mac, and cDCs revealing dominant IFN-1 contribution from myeloid cells followed by keratinocytes. 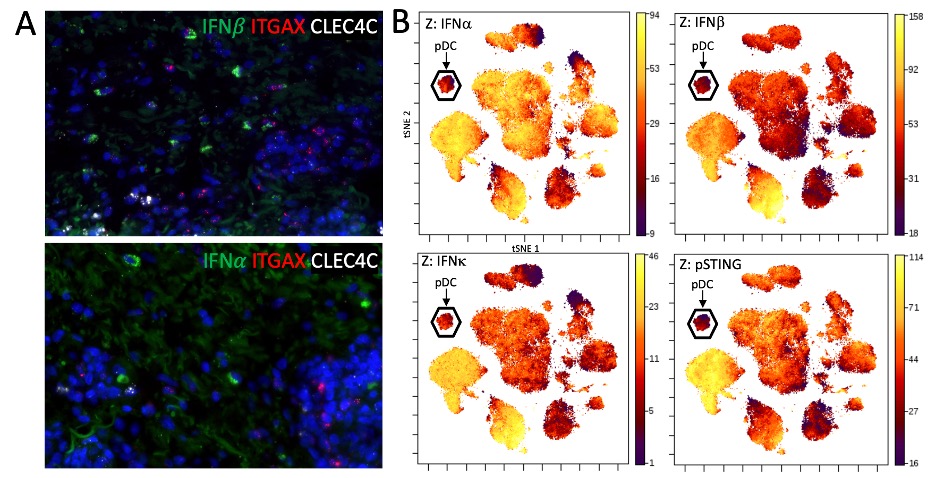 A) Representative images of mRNA in situ hybridization of IFN and IFN mRNA with markers for conventional dendritic cells (ITGAX) and plasmacytoid dendritic cells (CLEC4C). No observable overlap of type 1 interferon mRNA and CLEC4C was seen. B) tSNE plot of PBMCs from a CLE patient stained and acquired by CyTOF. Plasmacytoid dendritic cells, identified principally on CD123 expression, demonstrate little type 1 interferon staining as well as activation of the STING pathway (a signaling molecule upstream of type 1 interferon transcription) on the Z axis.
A) Representative images of mRNA in situ hybridization of IFN and IFN mRNA with markers for conventional dendritic cells (ITGAX) and plasmacytoid dendritic cells (CLEC4C). No observable overlap of type 1 interferon mRNA and CLEC4C was seen. B) tSNE plot of PBMCs from a CLE patient stained and acquired by CyTOF. Plasmacytoid dendritic cells, identified principally on CD123 expression, demonstrate little type 1 interferon staining as well as activation of the STING pathway (a signaling molecule upstream of type 1 interferon transcription) on the Z axis. 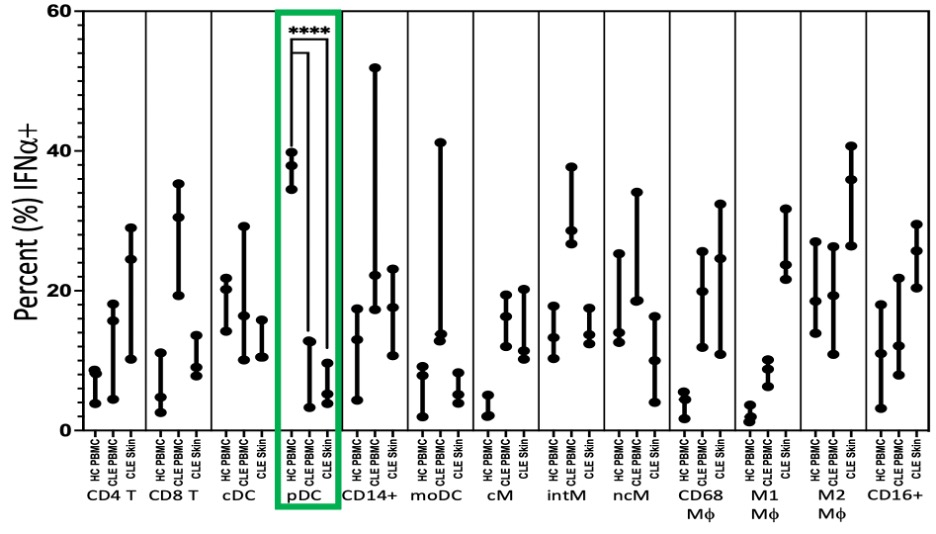 Flow cytometry of leukocytes eluted from CLE skin demonstrating significantly fewer IFN+ plasmacytoid dendritic cells (pDCs) in cutaneous lupus erythematosus (CLE) skin (Median 5.2%, p < 0.0001) and peripheral blood mononuclear cells (PBMCs) (12.7%, p < 0.0001) than healthy control PBMCs (37.9%). Abbreviations: cDC: classical dendritic cell; pDC: plasmacytoid dendritic cell; moDC: monocyte derived dendritic cell; cM: classical monocyte; intM: intermediate monocyte; ncM: nonclassical monocyte; Mf: macrophage.
Flow cytometry of leukocytes eluted from CLE skin demonstrating significantly fewer IFN+ plasmacytoid dendritic cells (pDCs) in cutaneous lupus erythematosus (CLE) skin (Median 5.2%, p < 0.0001) and peripheral blood mononuclear cells (PBMCs) (12.7%, p < 0.0001) than healthy control PBMCs (37.9%). Abbreviations: cDC: classical dendritic cell; pDC: plasmacytoid dendritic cell; moDC: monocyte derived dendritic cell; cM: classical monocyte; intM: intermediate monocyte; ncM: nonclassical monocyte; Mf: macrophage.