
Poster Session A
Rheumatoid arthritis (RA)
Kevin Cunningham, PhD
University of Minnesota
Minneapolis, MN, United States
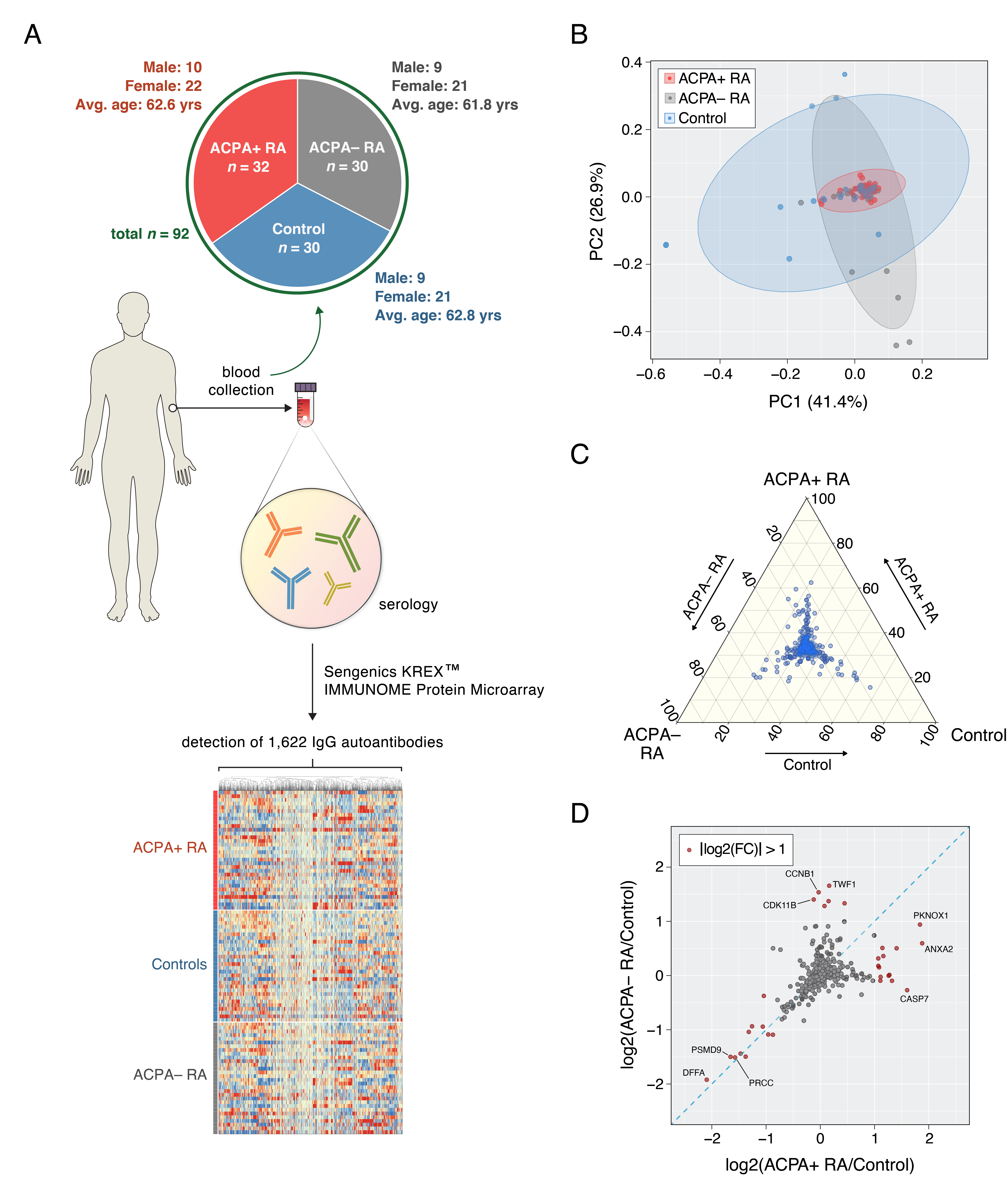 Figure 1. Study design overview and group-wise comparisons of autoantibody composition profiles. (A) Blood (serum) samples were collected to investigate serum autoantibody compositions in ACPA+ RA (n = 32), ACPA– RA (n = 30), and healthy controls (n = 30). Each serum sample was screened for 1,622 IgG autoantibodies using the Sengenics KREXTM IMMUNOME Protein Microarray. Raw relative fluorescence units (RFU) values representing serum autoantibody abundances were transformed using quantile normalization prior to all downstream analyses. Heatmap illustrates autoantibodies clustered according to abundance similarities across samples. (B) Principal component analysis on the 92 autoantibody profiles from ACPA+, ACPA–, and healthy controls. Controls showed the highest within-group heterogeneity in autoantibody composition whereas the ACPA+ RA group showed the least within-group heterogeneity. (C) Ternary plot showing mean abundances of 1,622 autoantibodies across ACPA+, ACPA–, and healthy controls. Coordinates of each point correspond to percentages of mean abundances and sum to 100. Despite most autoantibodies having similar mean abundances across the three study groups, several autoantibodies were found to have high abundance in one group (see points closer to the corners). (D) Fold-changes in mean abundances of 1,622 autoantibodies for the RA subgroups compared to the control group. x-axis and y-axis correspond to the fold-changes between ACPA+ RA and controls and between ACPA– RA and controls, respectively. Autoantibodies shown in red points have a fold-change of 2 (or greater) between groups. As observed in (C), several autoantibodies show fold-changes specific to an RA subgroup. The blue diagonal dashed line represents the line y=x.
Figure 1. Study design overview and group-wise comparisons of autoantibody composition profiles. (A) Blood (serum) samples were collected to investigate serum autoantibody compositions in ACPA+ RA (n = 32), ACPA– RA (n = 30), and healthy controls (n = 30). Each serum sample was screened for 1,622 IgG autoantibodies using the Sengenics KREXTM IMMUNOME Protein Microarray. Raw relative fluorescence units (RFU) values representing serum autoantibody abundances were transformed using quantile normalization prior to all downstream analyses. Heatmap illustrates autoantibodies clustered according to abundance similarities across samples. (B) Principal component analysis on the 92 autoantibody profiles from ACPA+, ACPA–, and healthy controls. Controls showed the highest within-group heterogeneity in autoantibody composition whereas the ACPA+ RA group showed the least within-group heterogeneity. (C) Ternary plot showing mean abundances of 1,622 autoantibodies across ACPA+, ACPA–, and healthy controls. Coordinates of each point correspond to percentages of mean abundances and sum to 100. Despite most autoantibodies having similar mean abundances across the three study groups, several autoantibodies were found to have high abundance in one group (see points closer to the corners). (D) Fold-changes in mean abundances of 1,622 autoantibodies for the RA subgroups compared to the control group. x-axis and y-axis correspond to the fold-changes between ACPA+ RA and controls and between ACPA– RA and controls, respectively. Autoantibodies shown in red points have a fold-change of 2 (or greater) between groups. As observed in (C), several autoantibodies show fold-changes specific to an RA subgroup. The blue diagonal dashed line represents the line y=x.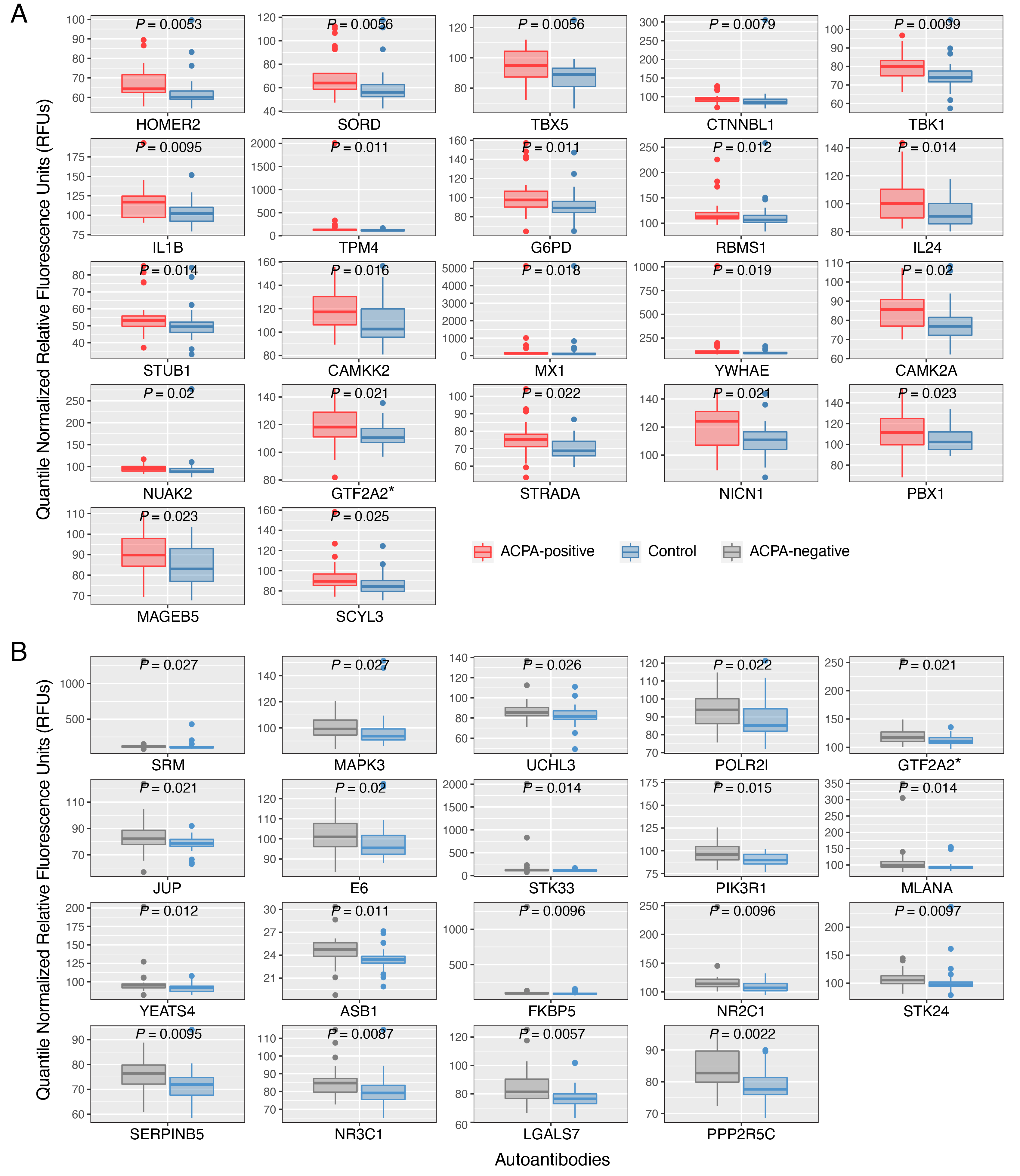 Figure 2. Autoantibodies identified to have significantly higher abundances in two subgroups of rheumatoid arthritis (ACPA+ RA and ACPA– RA) than in healthy controls. Patients with (A) ACPA+ RA and (B) ACPA– RA show higher abundances in 22 and 19 autoantibodies, respectively. Two-sided Mann–Whitney U test (P < 0.05) and Cliff’s Delta (|d| > 0.33) were used to find autoantibodies of significantly higher abundance. Standard box-and-whisker plots (e.g., center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range; points, outliers) are used to show numerical data. *, auto-antibody for GTF2A2 protein was found to have significantly higher abundances in both ACPA+ and ACPA– RA patient groups than in healthy controls.
Figure 2. Autoantibodies identified to have significantly higher abundances in two subgroups of rheumatoid arthritis (ACPA+ RA and ACPA– RA) than in healthy controls. Patients with (A) ACPA+ RA and (B) ACPA– RA show higher abundances in 22 and 19 autoantibodies, respectively. Two-sided Mann–Whitney U test (P < 0.05) and Cliff’s Delta (|d| > 0.33) were used to find autoantibodies of significantly higher abundance. Standard box-and-whisker plots (e.g., center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range; points, outliers) are used to show numerical data. *, auto-antibody for GTF2A2 protein was found to have significantly higher abundances in both ACPA+ and ACPA– RA patient groups than in healthy controls.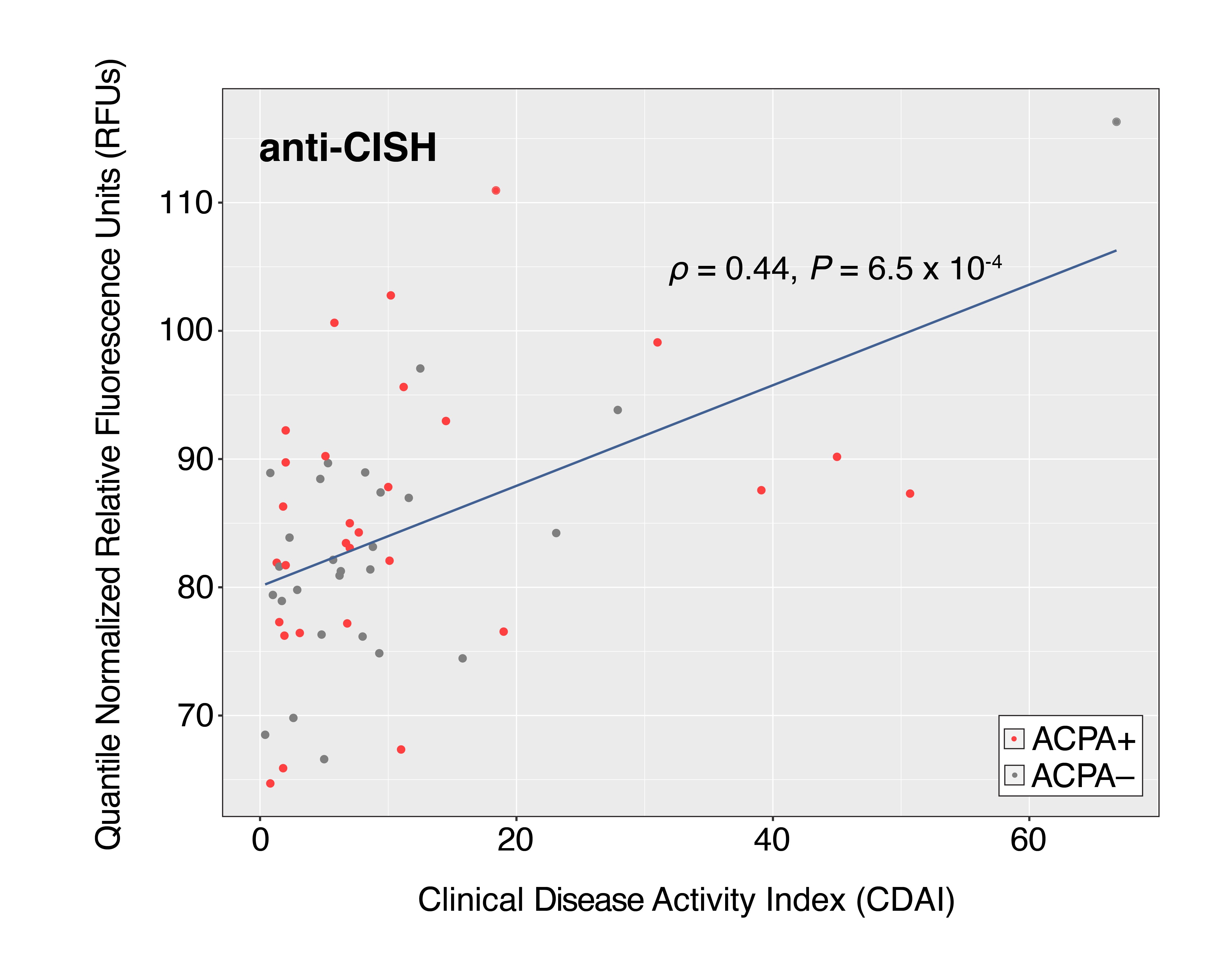 Figure 3. Autoantibodies targeting CISH proteins display the highest correlation with Clinical Disease Activity Index (CDAI). The blue line in the scatterplots represents the linear regression fit of Y~X, wherein X and Y correspond to CDAI and autoantibody abundance, respectively. The Spearman correlation coefficient ρ was used to measure the strength of the relationship between the two variables. Scatterplot contains 56 points corresponding to the number of RA patients whose CDAI scores were available.
Figure 3. Autoantibodies targeting CISH proteins display the highest correlation with Clinical Disease Activity Index (CDAI). The blue line in the scatterplots represents the linear regression fit of Y~X, wherein X and Y correspond to CDAI and autoantibody abundance, respectively. The Spearman correlation coefficient ρ was used to measure the strength of the relationship between the two variables. Scatterplot contains 56 points corresponding to the number of RA patients whose CDAI scores were available.